Hva påvirker når? Et livsløpsperspektiv på kognitiv aldring og hjernehelse
Kristine B. Walhovd1, 2 & Anders M. Fjell1, 2
1 Senter for livsløpsendringer i hjerne og kognisjon, Psykologisk institutt, Universitetet i Oslo
2 Senter for beregningsorientert radiologi og kunstig intelligens, Avdeling for radiologi og nukleærmedisin, Oslo universitetssykehus
Hva betyr egentlig «vellykket» kognitiv aldring? I media løftes ofte enkeltfenomener frem – som personer med høy funksjon eller tidlig demens, og det gis spesifikke livsstilsråd som er knyttet til å beholde høy funksjon. I denne artikkelen diskuterer vi hva forskning faktisk viser om kognitiv aldring, og hva som i større grad er myter enn realiteter. Vi oppsummerer kunnskapen om hvordan hjernen og kognitive funksjoner endrer seg gjennom livet, og viser at de store forskjellene man ser i hvordan eldre fungerer, ofte reflekterer stabile forskjeller som etableres tidlig, allerede i fosterliv og i løpet av barndom og ungdomstid, like mye som forskjeller i aldring i seg selv. Vi går deretter gjennom tre seiglivede påstander: at demensrisiko nesten kan halveres gjennom livsstilsendringer, at mye søvn er nødvendig for effektiv «hjernevask», og at høy utdanning bygger kognitiv reserve som beskytter mot aldersrelatert kognitiv svikt og demens. Til slutt diskuterer vi nye samfunnstrender som tilsier at risiko for kognitiv svikt og demens er avtagende, men advarer om at denne utviklingen er et resultat av gunstige samfunnsforhold som må vedlikeholdes. Skal den positive trenden fortsette, må vi investere i faktorer som bygger kognitiv kapasitet og motstandskraft gjennom hele livsløpet, særlig i de tidlige leveårene.
Nøkkelord: livsløpsperspektiv, utvikling, aldring, kognisjon, hjernehelse
What matters when?: A life-course perspective on cognitive ageing and brain health
What does ‘successful’ cognitive ageing actually mean? In the media, isolated phenomena are often highlighted, such as high-functioning individuals with or individuals with early-onset dementia, and specific lifestyle recommendations are frequently presented as key to maintaining cognitive performance. In this article, we discuss what research shows about cognitive ageing, and what is more myth than reality. We summarise current knowledge on how the brain and cognitive functions change across the lifespan, demonstrating that the large differences observed in the ways in which older adults function often reflect stable individual differences established early in life – already during foetal development and throughout childhood and adolescence – just as much as they reflect differences in the ageing process itself. We then examine three persistent claims: that dementia risk can be almost halved through lifestyle changes, that large amounts of sleep are necessary for effective ‘brain cleansing’, and that higher education builds cognitive reserve that protects against age-related cognitive decline and dementia. Finally, we discuss emerging societal trends suggesting that the risk of cognitive impairment and dementia may be decreasing, while cautioning that this development reflects favourable societal conditions that must be maintained. To sustain this positive trend, investment is needed in factors that build cognitive capacity and resilience across the entire lifespan, particularly in the early years of life.
Keywords: life-course perspective, development, ageing, cognition, brain health
De fleste kognitive funksjoner endrer seg når vi blir eldre, hjernehelsen og helsetilstanden i andre organsystemer endrer seg med dem. Men hva mener vi egentlig når vi snakker om «vellykket kognitiv aldring»? At en 80-åring fungerer som en gjennomsnittlig 40-åring? At man unngår demensdiagnose? Eller at man bevarer sitt eget funksjonsnivå så godt som mulig? En tilnærming har vært å studere eldre som fungerer spesielt godt – såkalte super-agers – eldre personer som skårer like høyt som langt yngre (Weintraub et al., 2025). Det kan for eksempel være 80-åringer som husker like godt, eller bedre, enn gjennomsnittet av middelaldrende. Slike personer kan være fascinerende, men de utgjør bare en liten andel av de eldre. De kan derfor lett gi en urealistisk standard for hva vi legger i «vellykket aldring».
Minst like viktig er et mer grunnleggende poeng: Å være en «super-ager» sier i utgangspunktet ingenting om hvordan man har endret seg, bare hvor man ligger an i forhold til andre. En 80-åring som skårer like høyt som unge voksne på en hukommelsestest, kan ha hatt ekstremt høye skårer som 30-åring, og faktisk ha tapt seg betydelig. Det illustrerer noe alle kliniske nevropsykologer vet, men som likevel (for) ofte glemmes i forskningen: Når vi bare ser på funksjonen her og nå, vet vi lite om hvordan personen har fungert tidligere.
En alternativ tilnærming legger derfor hovedvekten på faktisk endring over tid, slik vi ser i lengdesnittsstudier der deltagere følges med gjentatte undersøkelser. Nyberg og kolleger argumenterte allerede i 2012 for nytten av et brain maintenance-perspektiv («hjerne-bevaring») for å forstå hvordan hukommelse endrer seg i aldring, hvor vellykket aldring først og fremst forstås som liten eller ingen reduksjon (Nyberg et al., 2012), snarere enn et høyt nivå i seg selv. Når de samme personene følges gjentatte ganger, får man et annet bilde enn når man bare sammenligner ulike aldersgrupper: Noen opprettholder et relativt stabilt nivå av episodisk hukommelse over mange år også i høy alder, selv om gruppesnittet faller. Ut fra dette perspektivet kan en person som starter på et lavere kognitivt nivå, men holder seg bemerkelsesverdig stabil, i prinsippet ha en mer «vellykket» aldring enn en høytfungerende person som taper seg mye, selv om den siste fortsatt skårer høyest.
Så ja – det finnes eldre som fungerer bedre kognitivt enn de fleste 25-åringer, men det betyr ikke at de har unngått aldersendringer. Det er lite sannsynlig at noen opprettholder helt uendret hjernefunksjon gjennom et liv. De som er blant de beste som 80-åringer, har sannsynligvis vært blant de aller beste også som 30-åringer. Dette har minst to viktige implikasjoner: For å forstå kognitiv funksjon i aldring holder det ikke å studere bare eldre, og hvis vi vil bidra til å optimalisere kognitiv funksjon, må vi ta inn over oss at mange viktige innflytelser virker lenge før pensjonsalder. Vi må ha et livsløpsperspektiv på hjerne og kognisjon – og spørre hva som påvirker hvem når.
Hva skjer egentlig i hjernen i aldring?
For å forstå kognitiv aldring må vi se på de strukturelle og funksjonelle endringene i hjernen. De siste tiårenes forskning har vist at hjernen endres kontinuerlig gjennom hele livet, men at disse endringene er ujevnt fordelt. Det er ikke slik at hele hjernen krymper i samme takt. De største endringene ser vi i frontallappene (prefrontal cortex) og tinninglappene (temporal cortex). Dette er områder som er avgjørende for høyere kognitive funksjoner, og endringene her sammenfaller godt med det vi observerer kognitivt: Disse områdene er sentrale for planlegging, problemløsning, oppmerksomhet og episodisk hukommelse – altså nettopp funksjoner som oftest svekkes med alder. I kontrast er områder som primær synshjernebark ofte relativt sett bedre bevart. Strukturelt ser vi aldersendringer som tynnere hjernebark og noe redusert overflateareal, mindre volum i dypere strukturer som hippocampus og utvidelse av ventriklene (Figur 1). I tillegg endres trekk ved hjernens hvite substans, som utgjøres av nervefibrene og myelinet (isolasjonen) som sørger for rask og effektiv kommunikasjon mellom områder. Dette henger godt sammen med et av de mest robuste funnene i forskningen: at prosesseringshastighet og effektiv informasjonsflyt blir langsommere med alderen.
Figur 1
Hjernestrukturer gjennom livet
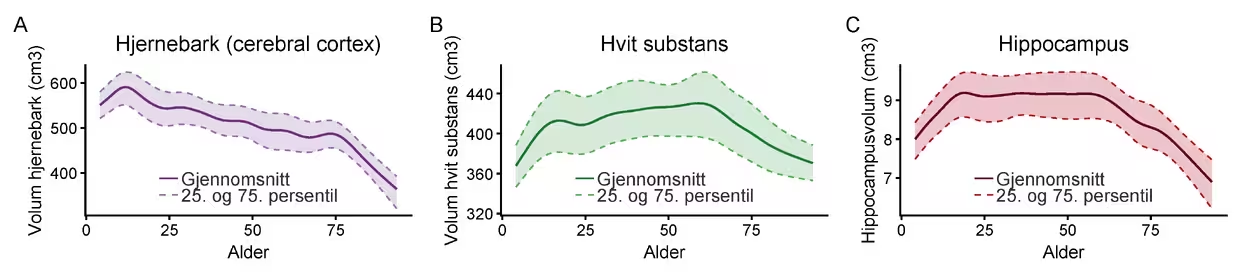
Merknad. Figuren viser volum av hjernebarken (grå substans) (A), hvit substans (B) og hippocampus (C) fra barndom til høy alder, basert på 4915 MRI-skanninger av 2242 kognitivt friske deltagere fulgt i opp til 16 år i studier ved LCBC, Universitetet i Oslo. Den heltrukne linjen viser gjennomsnittlig volum estimert med en kurvetilpasningsmodell. De stiplede linjene angir 25. og 75. persentil – halvparten av deltagerne faller på enhver alder innenfor dette båndet.
Men sammenhengen mellom hjernestruktur og kognitiv funksjon er mer kompleks enn «jo mer atrofi (vevstap), desto dårligere fungerer du». Selv om atrofi i hippocampus og midtre tinninglapper er knyttet til hukommelsessvikt (Vidal-Pineiro et al., 2025), forklarer strukturelle hjerneendringer ofte bare en moderat del av variasjonen i kognitiv endring hos ellers friske eldre. To personer kan ha omtrent samme grad av målbar hjerneatrofi, men likevel fungere svært ulikt kognitivt i hverdagen. Det samme ser vi når vi går fra hjernen til kognisjon (Figur 2). Kognitiv aldring er ikke én prosess, men et sett av ulike forløp. Såkalte «flytende» evner, som visuospatiale ferdigheter, prosesseringshastighet, arbeidsminne, fleksibel problemløsning og læring av nytt materiale, viser gjerne en gradvis svekkelse fra middelalderen og utover, selv om det er en del uenighet rundt på hvilken alder dette typisk er observerbart. Når det gjelder hukommelse, er episodisk hukommelse – minner om tid og sted – særlig sårbar. «Krystalliserte» evner, derimot, som verbal funksjon, allmennkunnskap og mange språklige ferdigheter, kan øke i voksen alder, og holder seg ofte relativt stabile langt inn i alderdommen. Et annet viktig poeng er at det er individuelle forskjeller i hvor mye kognitive funksjoner endrer seg med alder, også innenfor det vi kan kalle normalspekteret.
Figur 2
Kognitive evner gjennom livsløpet
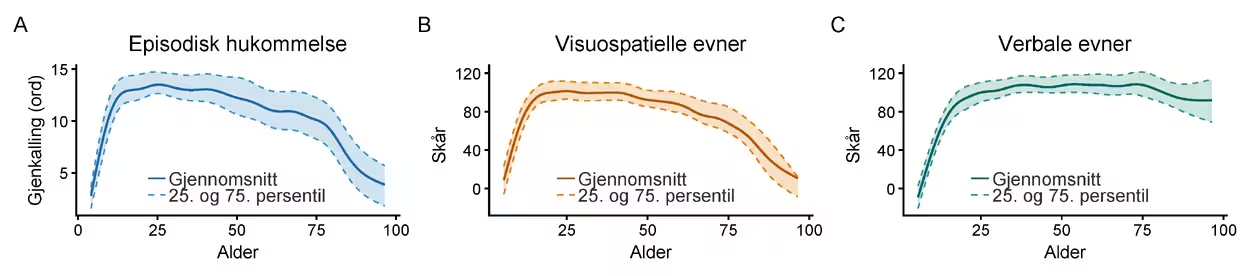
Merknad. Figuren viser tre mål på kognitiv funksjon fra barndom til høy alder. A: Episodisk hukommelse målt med gjenkalling av ordlister (CVLT), basert på 4774 undersøkelser av 2131 deltagere fulgt opp til 17,3 år i studier ved LCBC. B og C: Visuospatiale (terningmønster og matriseresonnering) og verbale (likheter og ordforråd) ferdigheter, begge fra WASI (Wechsler Abbreviated Scales of Intelligence), basert på henholdsvis 4121 og 4094 undersøkelser. Testene har et gjennomsnitt på 100 og standardavvik på 15, normert på deltagerne mellom 20 og 40 år. Alle skårer er korrigert for læringseffekter, estimert fra deltagere over 20 år for å unngå sammenblanding med kognitiv modning i barne- og ungdomsårene. Den heltrukne linjen viser gjennomsnittlig skår estimert med en kurvetilpasningsmodell. De stiplede linjene angir 25. og 75. persentil – halvparten av deltagerne faller på enhver alder innenfor dette båndet.
Dette leder til et tredje, ofte underkommunisert, poeng: Skillet mellom «normal» aldring og sykdom er langt mindre klart enn vi liker å tro. Obduksjonsstudier og moderne PET-avbildning viser for eksempel at en betydelig andel tilsynelatende kognitivt friske eldre har ansamlinger av typiske Alzheimer-markører, som amyloid og tau, i hjernen. Dette er det nylig også blitt mulig å måle med en blodprøve. Store studier tyder på at opptil 40 % av kognitivt friske 90-åringer har så høye nivåer av amyloid at de klassifiseres som «positive». Samtidig kan noen med relativt beskjedne mengder av slik patologi ha uttalte kognitive symptomer. Man kan altså ha en hjerne som ser «syk» ut, men fungerer helt normalt, og vice versa. Dette støtter et dimensjonalt syn på kognitiv aldring, hvor overgangen fra normalitet til patologi ofte ikke er klart avgrenset, men går langs et kontinuum. Dette gjelder spesielt i høy alder, der forskjellene mellom en som er frisk, og en med en Alzheimer-diagnose kan være mye mindre enn mellom en frisk og en dement 60-åring. Dette åpner for et mer nyansert syn: Hjernen eldes, men hvordan dette faktisk slår ut kognitivt, er resultatet av en rekke faktorer som kan virke gjennom et langt liv.
Figur 3
Individuelle forløp for hippocampusvolum og episodisk hukommelse
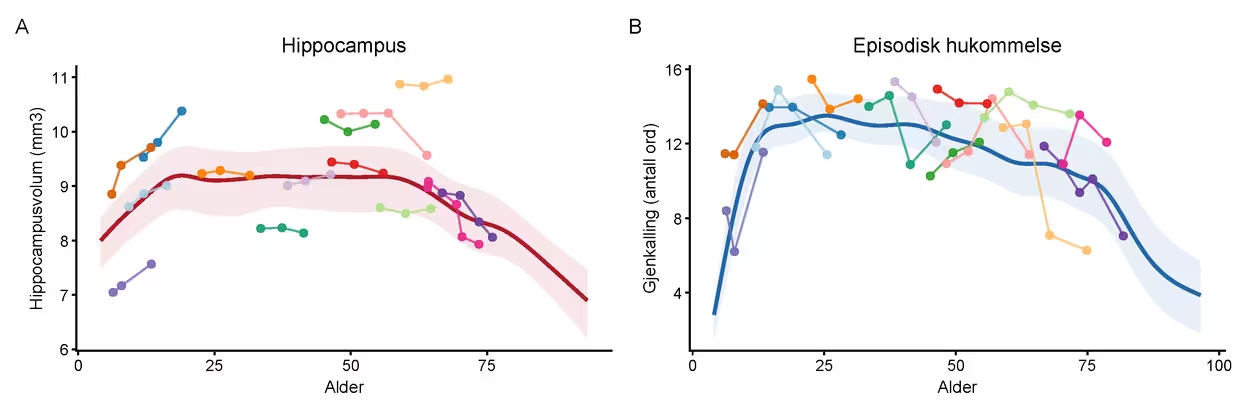
Merknad. Figuren illustrerer individuelle forskjeller i hjernestruktur og kognitiv funksjon over tid. Bakgrunnskurven viser gruppegjennomsnittet med 25. og 75. prosentilbånd (se Figur 1 og 2 for detaljer). De fargede linjene og punktene representerer 14 tilfeldig utplukkede enkeltdeltagere med minst tre målinger. Samme farge angir samme person i begge paneler. Som man kan se, er det store forskjeller både i hvor høyt deltagerne ligger, og hvor mye de endrer seg over tid. Hvis man studerer fargene nøye, er det lett å finne deltagere som viser en mer positiv utvikling i hippocampus enn på hukommelsestesten, og omvendt, noe som viser at det er flere ting som avgjør hvor godt man fungerer.
Å fremme god hjernehelse
Hva kan gjøres for å fremme høy kognitiv funksjon og god hjernehelse? For å forstå hva som gir god hjernehelse i eldre år, må vi først erkjenne en fundamental, men ofte oversett dynamikk: Grunnlaget for hvor godt hjernen fungerer som 80-åring, legges i stor grad før vi fyller 20. Selv om vi i aldringsforskningen ofte leter etter årsaker til kognitiv svikt sent i livet, viser livsløpsdata at individuelle forskjeller i hjernestruktur og kognitiv funksjon er bemerkelsesverdig stabile gjennom hele livet. Et av de mest overbevisende funnene kommer fra Lothian Birth Cohort-studiene i Skottland, ledet av Ian Deary. En stor gruppe barn som tok en intelligenstest som 11-åringer, ble testet igjen i 70-, 80- og 90-årsalderen. Resultatene viser en forbløffende høy korrelasjon (rundt.70) mellom prestasjonene i barndommen og i alderdommen (Deary et al., 2013). Det betyr at selv om den kognitive funksjonen svekkes hos alle gjennom livsløpet, endres vi i stor grad i samme takt. De som skåret høyt som barn, skåret som regel høyt som gamle. Likevel var det liten sammenheng mellom hvor høyt deltagerne skåret i barndommen, og hvor mye de endret seg i 80-årene. Det er altså ikke så lett å bytte plass i køen; rangordningen mellom individer er bemerkelsesverdig stabil.
Dette mønsteret bekreftes også når vi ser direkte på hjernestruktur. I en nylig studie fra vår egen forskningsgruppe analyserte vi hvordan ulike hjernestrukturer endret seg over tid gjennom voksenlivet (Grodem et al., 2025). Funnene viser at individuelle forskjeller i hjernevolum domineres av stabile forskjeller, altså det nivået man tar med seg inn i voksenlivet, helt frem til høy alder. Forskjeller i hvor raskt hjernen eldes (atrofiraten), begynte først å få betydning for rangordningen mellom folk når de nærmet seg 80 år. Implikasjonen er tydelig: Hvis vi ønsker å forbedre hjernehelse og redusere forekomsten av kognitiv svikt, ligger det et stort potensial i å løfte utgangspunktet. Mye tyder på at det er vanskeligere å bremse aldringsprosessen enn det er å sørge for at flest mulig starter alderdommen med et best mulig utgangspunkt.
Tidlige år former hjernen for livet
Når vi diskuterer hvordan vi kan opprettholde god kognitiv funksjon og hjernehelse i eldre år, er det lett å sette søkelys på de som er, eller snart er, eldre nå. Men forskning tyder altså på at mye av grunnlaget for hvor godt hjernen vår fungerer som 80-åringer, ble lagt i utvikling tidlig i livet (Walhovd et al., 2023). En gjennomgang av livsløpsdata viser at mye av forskjellene vi ser mellom eldre mennesker, egentlig er «gammel variasjon», og at faktorer som virker før fødselen og i tidlig barndom, har minst like stor betydning for hjernens struktur i alderdommen som selve aldringsprosessen. For eksempel viser studier at fødselsvekt, som er en indikator på utvikling i fosterlivet, henger sammen med hjernestruktur og kognitiv funksjon gjennom hele livsløpet, helt opp i 80-årsalderen. Ved å sammenligne norske barn ned til 4 år med voksne og eldre ser vi at områdene i hjernebarken der strukturelle trekk er forbundet med generell kognitiv funksjon, overlapper i svært stor grad på tvers av livsløpet, og hjerneendringene over tid er omtrent like for de som skårer høyt, og de som skårer lavere (Walhovd et al., 2016). Dette indikerer at de nevrale forutsetningene for kognitiv kapasitet etableres tidlig i utviklingen og vedvarer som stabile trekk gjennom livet, snarere enn primært å være et resultat av hvor godt hjernen motstår aldring. Genetikk spiller også en avgjørende rolle her; tvillingstudier indikerer at arvelighet forklarer en betydelig del av variasjonen i både hjernestruktur og kognitiv endring, og at genetiske faktorer bidrar til stabilitet over tid (Fjell et al., 2015).
Det er også en utbredt antakelse innen aldringsforskning at hjernen blir mer sårbar for miljøpåvirkning jo eldre vi blir. Men empirien peker ofte motsatt vei: Hjernen ser ut til å være mest plastisk og mottakelig for påvirkning tidlig i utviklingen. For eksempel ser vi at flere risikofaktorer som trekkes frem som viktige livsstilsfaktorer midt og sent i livet, for eksempel rusmiddelpåvirkning, har en betydelig større påvirkning på hjernen når de forekommer i fosterlivet og tidlig barndom (Walhovd et al., 2023). Konsekvensen av dette er at det vi ofte tolker som «vellykket aldring», at noen har bevart god funksjon og hjernehelse, i stor grad reflekterer et godt utgangspunkt etablert tidlig i livet. Ved å ignorere disse tidlige faktorene risikerer vi å overvurdere betydningen av livsstil og helsevalg i alderdommen. Det betyr ikke at det vi gjør sent i livet, er uviktig, men at effekten av tiltak rettet mot barn og unge, som å sikre god prenatal omsorg, oppvekst og utdanning, sannsynligvis har en større og mer varig effekt på befolkningens hjernehelse enn noe vi kan gjøre i livets senere faser.
Hva skjer senere?
I avsnittene over har vi argumentert for at stabile forskjeller mellom mennesker etableres tidlig i livet, og at disse forskjellene fortsatt er viktige i høy alder. Det betyr likevel ikke at alle eldes likt. Vi ser tydelige individuelle forskjeller i hvordan for eksempel hukommelse endrer seg med alder (Fjell et al., 2025), noe som blant annet henger sammen med hvor store hjerneendringer den enkelte gjennomgår (Vidal-Pineiro et al., 2025). Mange faktorer bidrar til slike endringer, men kanskje er det viktigste spørsmålet ikke hva som gjør oss like, men hva som gjør oss forskjellige: Hva forklarer at noen eldes raskere enn andre? Noe kan vi ikke gjøre noe med, som genetiske forskjeller. For eksempel har bærere av APOE ε4-allelen, den sterkeste kjente risikofaktoren for sporadisk Alzheimers sykdom, i gjennomsnitt noe brattere aldringsforløp i hjernen (Roe et al., 2024). Samtidig ser vi at demensinsidensen i flere land er fallende: Selv om antallet personer med demens øker på grunn av en eldre befolkning, er sannsynligheten for å få demens på en gitt alder lavere nå enn tidligere. En stor metaanalyse fra Europa og USA har anslått rundt 13 % reduksjon i risiko per tiår (Wolters et al., 2020). Det er usannsynlig at genetikk kan forklare dette, og slike funn brukes ofte som argument for at modifiserbare faktorer, altså livsløpsfaktorer vi i prinsippet kan påvirke, spiller en rolle for kognitiv funksjon i aldring.
Her er det likevel viktig å ha tunga rett i munnen. Vi vet fortsatt ikke om den lavere demensrisikoen skyldes at folk kommer inn i alderdommen med høyere kognitivt nivå, for eksempel på grunn av Flynn-effekten. Flynn-effekten viser til at man ser i gjennomsnitt høyere kognitiv prestasjon over tid for nyere fødselskohorter. Tidsforløpet varierer noe over land, men i Norge så vi økende kognitiv skårer i ung voksen alder for mennesker født på 1900-tallet, helt frem til kohortene født på midten av 70-tallet. Dagens eldre kan dermed ha «mer å gå på» før de krysser en klinisk terskel for demens enn det deres foreldre hadde. Norske data viser at lav skår på intelligenstester ved 18-årsalder (sesjonsdata) omtrent dobler risikoen for tidlig demens (Bratsberg et al., 2024), og dette tyder på at slike mekanismer kan være viktige: Generasjonsforskjeller i kognitiv funksjon tidlig i livet kan bidra til lavere demensrater flere tiår senere. Samtidig er det nyere data som tyder på at dagens eldre faktisk har mindre kognitiv nedgang over tid enn tidligere kohorter. Nøyaktig hva som driver dette, er vanskelig å fastslå sikkert, men det finnes mange kandidater, som bedrede levekår generelt, inkludert for eksempel redusert eksponering for miljøforurensning, mindre røyking og bedre tilgang til forebyggende behandling for hjerte- og karsykdommer.
Når det gjelder hva enkeltpersoner kan gjøre i middelalder og alderdom for å påvirke kognitiv aldring, er bildet mer usikkert. Å identifisere sterke, kausale mekanismer for endring på individnivå er betydelig vanskeligere enn å påvise sammenhenger, men vi vil trekke frem tre faktorer der det er overbevisende evidens for faktisk påvirkning på kognitiv funksjon:
Målrettet kognitiv trening: Mange studier viser at målrettet kognitiv trening kan gi forbedringer på de oppgavene man trener på, og noen ganger ser man også målbare endringer i hjernen (Engvig et al., 2010). Evidensen for bred overføring, at treningen gir varige gevinster i helt andre typer oppgaver eller i daglig fungering, er derimot liten. Effekten ser ofte ut til å være spesifikk og moderat. Man blir kun bedre på akkurat det man trener på.
Tiltak som bedrer psykisk helse og livskvalitet: Tiltak som reduserer depresjon, øker sosial aktivitet og gir opplevelse av mestring og glede, kan ha positive effekter på både subjektiv kognitiv funksjon og livskvalitet. Selv om det ikke enkelt lar seg dokumentere at de reduserer demensrisikoen i seg selv, er dette sannsynligvis blant de viktigste og mest håndfaste tingene man kan gjøre for å ha det bra med den hjernen man har. Dessuten husker de fleste mindre når man er engstelig, så for eksempel å engste seg for demens kan paradoksalt nok føre til at man husker dårligere.
«Det som er bra for hjertet, er ofte bra for hjernen»: Fysisk aktivitet, blodtrykkskontroll, god behandling av diabetes og generelt kardiovaskulær risikoreduksjon er gjennomgående forbundet med bedre kognitiv funksjon og lavere demensrisiko på gruppenivå. Likevel er det sjelden vi kan si med sikkerhet hva slags effekt et enkelt tiltak vil ha for én bestemt person, effekter vil antagelig akkumuleres over tid, og det er usikkert om slike tiltak har umiddelbar effekt på hjernehelse. Evidensen er sterkest på befolkningsnivå, men det er grunn til å tro at for folk i risikogrupper kan disse faktorene ha en viss betydning. Man bør imidlertid se slike tiltak som et samfunnsmål, der det er snakk om at man over tid kanskje kan løfte gjennomsnittshelsen i befolkningen med en viss gunstig effekt også på kognitiv funksjon. Dette kan imidlertid ikke tolkes som at den enkelte har sin skjebne i egne hender, eller selv i stor grad kan påvirke egen kognitiv funksjon i aldring.
Oppsummert: Det finnes flere ting som sannsynligvis kan bidra positivt til kognitiv aldring, men vi har fortsatt ikke grunnlag for å love at spesifikke tiltak forhindrer eller utsetter demens hos enkeltpersoner. De beste rådene handler derfor om det samme som for fysisk helse generelt: Det bør være et mål å skape samfunn der den enkelte kan utvikle seg til den beste versjonen av seg selv, og der det optimaliseres for hjerte- og karhelse, at folk kan holde seg mentalt og sosialt aktive, og leve liv som gir mening og trivsel, samtidig som vi erkjenner at biologisk aldring og genetiske faktorer setter rammer vi ikke kan gjøre noe med. I det følgende vil vi gå gjennom tre utbredte oppfatninger – det finnes mange flere – og gi en kritisk vurdering av hvor solid kunnskapsgrunnlaget for hver av dem faktisk er.
Tre myter om kognitiv aldring
Du kan redusere demensrisiko med 45 %
Denne påstanden stammer fra Lancet Commission on dementia prevention, intervention, and care (Livingston et al., 2024), som har anslått at rundt 45 % av demenstilfellene i befolkningen potensielt kan forebygges ved å redusere 14 risikofaktorer. I pressen formuleres dette ofte som om det gjelder deg som enkeltperson. Det høres ut som et konkret løfte – «gjør du alt riktig, halverer du nesten risikoen» – men tallene er verken individuelle, kausale eller spesielt presise.
For det første er 45 %-tallet et populasjonsmål, ikke en individuell effektstørrelse. Det sier – under svært sterke forutsetninger – noe om hvor stor andel av tilfellene i en hel befolkning som kunne vært unngått dersom alle risikofaktorer både var fullt ut årsaker og helt eliminert. I praksis er mange av faktorene tett sammenvevd (for eksempel utdanning, sosial deltagelse og genetisk pleiotropi [at de samme genene påvirker flere ting]), og beregningen forutsetter at det ikke finnes underliggende faktorer som påvirker flere av dem samtidig, noe vi vet ikke stemmer. Kommisjonen understreker selv at estimatene er usikre og sannsynligvis overvurderer effekten, men det er sjelden det som havner i overskriftene. I tillegg forutsetter 45 %-tallet tilstedeværelsen av alle risikofaktorene. I Norge er det for eksempel under 2 % som drikker mer enn 21 alkoholenheter i uken («excessive alcohol consumption»), rundt 7 % som røyker daglig, rundt 6 % som har diabetes, og kanskje rundt 10 % som har hatt traumatisk hjerneskade. For de fleste enkeltpersoner vil den teoretiske «maksgevinsten» derfor være en brøkdel av 45 %, rett og slett fordi de mangler disse risikofaktorene.
For det andre bygger anslagene i stor grad på observasjonsstudier, altså sammenhenger der man, i beste fall, følger folk over tid, men ikke kontrollerer den kausale effekten av hver enkelt faktor. Slike data er viktige, men det er svært vanskelig å skille årsak fra virkning. Nedsatt hørsel, depresjon eller fysisk inaktivitet kan for eksempel være tidlige symptomer på eller konsekvenser av den samme sykdomsprosessen som senere gir demens, heller enn selvstendige årsaker. Når man så oversetter slike sammenhenger til «prosent redusert risiko» og media presenterer dem som om de gjelder enkeltpersoner, gir det et skinn av kausal presisjon som ikke finnes i datagrunnlaget. Nyere forsøk på å komme nærmere kausalitet illustrerer problemet. Hørselstap er en enkeltfaktor som Lancet-kommisjonen anslår har blant de sterkeste effektene (7,0 %). ACHIEVE-studien, den første store randomiserte studien som undersøkte om høreapparat kan bremse kognitiv nedgang, fant ingen samlet effekt på generell kognitiv funksjon (Lin et al., 2023). I en forhåndsdefinert høyrisikogruppe med økt kardiovaskulær risiko så man en liten fordel for intervensjonsgruppen på generelt kognitivt nivå (β ≈ 0,19 SD (standardavvik) over 3 år, 95 % KI (konfidensintervall) 0,02–0,36; p=0,027), tilsvarende rundt 3 «IQ-poeng» over tre år på en skala med gjennomsnitt 100 og standardavvik 15. Andre kognitive domener, inkludert hukommelse, som typisk er mest sensitivt for tidlig demens, viste ingen effekt. Det positive resultatet kom fra en sensitivitetsanalyse med flere sammenligninger, og gjelder endring i testskårer, ikke forekomst av demens. Samlet sett gir ACHIEVE derfor støtte til at hørselstiltak kanskje kan ha en begrenset gunstig effekt for enkelte risikogrupper, men langt fra en sterk, generell dokumentasjon for at hørselstiltak i seg selv forebygger demens i befolkningen.
Et lignende bilde ser vi i FINGER-studien (Finnish Geriatric Intervention Study to Prevent Cognitive Impairment and Disability). Den første FINGER-studien var en toårig, bredt anlagt livsstilsintervensjon for eldre med forhøyet demensrisiko, som kombinerte kostholdsråd, fysisk aktivitet, kognitiv trening og oppfølging av vaskulær risiko (Ngandu et al., 2015). Studien trekkes ofte frem som et hovedargument for at livsstilsendringer kan forebygge kognitiv svikt. FINGER viste statistisk signifikante, men svært små forbedringer i kognitiv funksjon etter to år: Forskjellen i endring mellom intervensjons- og kontrollgruppen var på rundt 0,022 standardavvik på et sammensatt kognitivt mål, tilsvarende om lag en tredels «IQ-poeng». Dette er forenlig med at omfattende livsstilstiltak kan ha en gunstig effekt på kognisjon hos utvalgte risikogrupper i befolkningen, men ligger svært langt unna å dokumentere noe i nærheten av 45 % reduksjon i demensrisiko for enkeltpersoner. I tillegg var deltagerne en selektert gruppe eldre med økt vaskulær risiko, slik at resultatene i begrenset grad kan generaliseres til den brede eldre befolkningen.
Et tredje problem er overgangen fra populasjon til individ, og dette problemet består selv om vi ser bort fra alle innvendingene ovenfor. La oss for argumentets skyld anta at alle metodologiske problemer er løst: at vi hadde presise, kausale effektestimater for hver risikofaktor, og at disse virkelig lot seg summere til 45 %. Selv da ville det ikke følge at en konkret person kan redusere sin individuelle risiko med 45 % ved å endre livsstil. Grunnen er at gjennomsnittlige effekter på tvers av en befolkning skjuler enorm variasjon mellom enkeltpersoner. En persons grunnrisiko for demens er i betydelig grad bestemt av forhold som genetisk sårbarhet og tidlige livsbetingelser, faktorer som ikke nødvendigvis lett lar seg påvirke i voksen alder, og som avgjør hvor mye rom det er for forbedring. For en person med lav grunnrisiko kan gevinsten av livsstilsendringer være neglisjerbar; for en annen med høy grunnrisiko kan den samme endringen bety mer, men bare hvis livsstilsendringen påvirker nettopp de risikofaktorene denne personen har. De statistiske estimatene sier noe om gjennomsnittseffekter under idealiserte forutsetninger, ikke om skjebnen til en konkret 75-åring som snører på seg joggeskoene.
Til slutt er Lancet-kommisjonens rammeverk ikke en reell livsløpsmodell, selv om den ofte omtales som det. Vi vet at mange faktorer har langt større innvirkning på hjernen når de virker tidlig i livet, men dette tas i liten grad hensyn til. Den eneste «tidliglivsfaktoren» er «less education», og i 2024-oppdateringen er det ikke engang tydelig definert hva dette innebærer. Andre forhold vi vet er svært viktige for kognitiv funksjon i aldring – som generasjonsforskjeller i skole, ernæring, helse og kognitiv stimulering, som antagelig bidrar til Flynn-effekten, er ikke inkludert. Flynn-effekten har vært betydelig, rundt 3 IQ-poeng per tiår i store deler av det 20. århundre (Pietschnig & Voracek, 2015), og det er vanskelig å se hvordan man kan diskutere forebygging av demens uten å ta slike strukturelle forbedringer med i bildet. I tillegg gir det lite mening at enkelte faktorer klassifiseres som «senlivsrisiko» (f.eks. luftforurensning og sosial isolasjon), mens andre kun betraktes som «midtlivsrisiko» (f.eks. depresjon og hørselstap), når det ikke finnes gode grunner til at de ikke også skulle være viktige tidligere i livet.
Alt dette betyr ikke at livsstil er irrelevant. Kontroll av blodtrykk, fysisk aktivitet og god psykisk helse har som nevnt veldokumenterte fordeler for hjerte-karsystemet, funksjon og livskvalitet, og sannsynligvis også for kognisjon hos mange. Poenget er at påstanden om «45 % reduksjon i demensrisiko» i beste fall er et grovt, optimistisk anslag på hvor stor del av sykdomsbyrden som kanskje kan påvirkes på befolkningsnivå gjennom langvarige strukturelle og adferdsmessige endringer. I verste fall gir den et urealistisk løfte og et urimelig press på enkeltmennesker, og risikerer å overskygge den betydelige rollen som tidlige livsbetingelser, genetisk sårbarhet og tilfeldigheter vi ikke har kontroll over, spiller i kognitiv aldring. En frisk hjerne er en hjerne som søker forsterkning, noe som oppleves som belønnende. Det varierer i hvilken grad vi opplever antatte risikofaktorer som forsterkende eller forstyrrende, som for eksempel alkoholbruk, grad av fysisk aktivitet eller kosthold. Oppsummert mener vi evidensen ikke tilsier at enkeltpersoner bør gjøre svært store inngrep i eget liv for å minimere demensrisiko dersom dette oppleves som veldig negativt. Å løpe hver dag og kun spise knekkebrød til lunsj er noe å anbefale kun for de som liker det. Det er etter vårt syn lite sannsynlig at tid investert i noe man opplever som negativt, kan settes på en konto der man får igjen god og morsom tid med renter senere i livet. Derimot er det viktig for god helse å oppleve livet meningsfylt her og nå og å være glad.
God «hjernevask» krever mye søvn
En av de mest populære teoriene er at søvnens hovedoppgave er å «rense» hjernen for metabolske avfallsstoffer, som Alzheimer-relatert amyloid, via det såkalte glymfatiske systemet. Tanken er enkel og intuitiv: Sover du for lite, hoper avfallsstoffene seg opp, og hjernen brytes ned. Dette har bidratt til en utbredt forestilling om at vi må sove mye for at hjernen skal holde seg frisk. Ser vi systematisk på forskningen hos mennesker, er imidlertid evidensen for en slik direkte årsakskjede overraskende svak. Hvis lang søvn var kritisk for å bevare hjernen, skulle vi forvente at de som sover lite, jevnt over har mer hjerneatrofi og kognitiv svikt i aldring, og de som sover mye, mer bevarte hjerner. Dette er ikke det mønsteret vi ser. Tvert imot finner flere studier som følger deltagerne over tid, ingen sterk sammenheng mellom søvnlengde og endringer i hjernestruktur, som hippocampusatrofi eller fortynning av hjernebarken (Fjell et al., 2023). Når det først dukker opp sammenhenger, er de ofte små, inkonsistente og forsvinner typisk når man kontrollerer for andre faktorer. Tilsvarende ser man heller ikke at mye søvn systematisk beskytter hukommelse eller annen kognitiv funksjon hos ellers friske voksne.
Det kanskje tydeligste tegnet på at «mer er bedre»-hypotesen ikke holder, er funnene om lang søvn. Det finnes lite belegg for at det å sove mer enn rundt syv timer per natt i seg selv beskytter hjernen. Tvert imot har flere store studier funnet at lang søvn er assosiert med dårligere kognitiv funksjon og økt risiko for kognitiv svikt og demens. Det undergraver ideen om søvn som en enkel dose–respons-kur, der flere timer gir renere og friskere hjerne. Et mer sannsynlig mønster er at endringer i søvnmønster ofte er symptomer på underliggende prosesser, som begynnende nevrodegenerasjon, depresjon eller somatiske sykdommer.
Teorien om «hjernevask» støter også på problemer når vi tar et evolusjonært perspektiv. Gjennom dyreriket er det stor variasjon i hvor mye søvn ulike arter og individer trenger, uten noen enkel kobling mellom søvnlengde og kognitiv kapasitet. Mennesker sover faktisk kortere enn mange andre primater, til tross for at vi har større hjerner og mer avanserte kognitive funksjoner. Dersom lang søvn var helt avgjørende for å rense en stor hjerne, skulle vi forventet at mennesker sov mer, ikke mindre, enn sjimpanser og bavianer. At vi har utviklet oss til å være «kort-sovere», tyder på at søvnens funksjon i stor grad handler om effektivitet og fleksibilitet, et system som kan tilpasses miljøet, heller enn en passiv renseprosess som krever stadig flere timer. Faktisk er søvn en svært plastisk egenskap som varierer mye med kortvarige eller langvarige skifter i omgivelsene, og det ser ikke ut til å eksistere noen deterministisk «ideell søvnlengde» (Fjell & Walhovd, 2024).
Alt dette betyr ikke at søvn er uviktig for hjernen. Akutt søvnmangel kan påvirke oppmerksomhet, humør og kognitiv prestasjon, og langvarige søvnproblemer henger sammen med redusert psykisk helse. Men bildet som ofte formidles, at kort søvn i seg selv er en driver av nevrodegenerasjon hos ellers friske mennesker, har ikke empirisk grunnlag. Det er ingen grunn til å tro at man kan sove seg til en lykkelig alderdom eller en demensfri hjerne.
Høy utdanning beskytter mot demens
En utbredt oppfatning er at høy utdanning beskytter mot demens ved å bygge opp en såkalt kognitiv reserve. I forskningslitteraturen brukes begrepet kognitiv reserve til å beskrive at noen mennesker kan ha tilsynelatende normal kognitiv fungering selv om hjernen viser betydelig aldring eller sykdom. Tanken har vært at høyere utdanning gjør hjernen mer motstandsdyktig, at år på skolebenken aktivt styrker nettverk og strategier som gjør at man tåler mer sykdomsforandringer eller vevstap før symptomene viser seg. Går vi empirien etter i sømmene, må dette bildet nyanseres kraftig. Det er godt dokumentert at skolegang gjør at vi presterer bedre på kognitive tester. En stor metaanalyse av naturlige eksperimenter, som reformer innført til ulike tider, viser at hvert ekstra år på skole gir i størrelsesorden 1–5 IQ-poeng økning, og at effekten varer livet ut (Ritchie & Tucker-Drob, 2018). Mer skolegang gjør at man kan gå inn i voksenliv og alderdom med et høyere kognitivt nivå enn man ellers ville gjort. Spørsmålet er om dette også betyr at utdanning bremser aldringen eller reduserer sårbarheten for demens.
Nyere data tyder på at svaret i stor grad er nei. I en stor norsk studie av over 200 000 menn ble sammenhengen mellom utdanning og risiko for tidlig demens undersøkt. Som forventet var lav utdanning forbundet med høyere demensrisiko, men denne sammenhengen forsvant nesten helt når man kontrollerte for kognitiv funksjon ved militær sesjon rundt 18-årsalderen (Bratsberg et al., 2024). Dette tyder på at det er det kognitive funksjonsnivået etablert tidlig i livet, ikke antall år med utdanning i seg selv, som er den sentrale risikofaktoren. Utdanning kan bidra til å øke skårene, men når man først har tatt høyde for hvor godt folk faktisk fungerer i ung voksen alder, gir ekstra utdanning liten eller ingen ytterligere beskyttelse mot tidlig demens.
Også når vi ser på forløp over tid, er bildet det samme: Analyser av livsløpsdata viser at utdanning ikke ser ut til å fungere som en «brems» på kognitiv aldring (Fjell et al., 2025). Høyt og lavt utdannede taper seg i gjennomsnitt omtrent like raskt, selv om utgangspunktet er forskjellig. Det finnes heller ikke sterke holdepunkter for at høy akademisk utdanning gir bedre beskyttelse mot aldersrelaterte endringer i hjernehelse enn det å ha fullført videregående. Den store folkehelsegevinsten ser ut til å ligge i at flere gjennomfører grunnskole og videregående, som løfter hele befolkningens kognitive nivå, ikke i at flere tar svært lang utdanning. At personer med høy utdanning statistisk sett får demensdiagnose senere, betyr derfor sannsynligvis først og fremst at de, av grunner som ikke nødvendigvis har med utdanning i seg selv å gjøre, starter høyere og dermed har en lengre vei å falle før de passerer en diagnostisk terskel, ikke at utdanningen har gitt dem en magisk «reserve» de kan bruke for å beskytte seg mot aldersrelaterte endringer i hjernen eller andre sykdomsprosesser.
Veien videre
Vi ser for tiden en betydelig optimisme knyttet til forebygging av aldersrelatert kognitiv svikt og demens, som reflektert i Lancet-kommisjonens anslag, og data fra Europa og USA tyder på at risikoen for demens er redusert med rundt 13 % per tiår i senere fødselskohorter. Det betyr at selv om antallet personer med demens vil øke fordi det blir flere eldre, er sannsynligheten for å få demens ved en gitt alder lavere nå enn den var for besteforeldregenerasjonen.
Spørsmålet er hvorfor vi ser denne positive utviklingen. Det er klart at senere generasjoner har et høyere kognitivt utgangsnivå, sannsynligvis i stor grad på grunn av forhold som virket tidlig i livet, som bedre ernæring, helse, skole og oppvekstsvilkår. Når det gjeldet spørsmålet om nyere kohorter også har en gunstigere aldringstakt, med mer stabile kognitive forløp i eldre år, er forskningen mer sprikende. Flere studier finner at dagens eldre ikke endrer seg mindre kognitivt enn deres foreldre gjorde på samme alder, mens andre resultater tyder på det motsatte. Årsakene vil i så fall være andre enn de som forårsaker det høyere utgangsnivået. Hvis den finnes, vil en slik «stabilitetsgevinst» trolig være drevet av brede samfunnsendringer som akkumuleres gjennom livsløpet, som bedre kontroll av kardiovaskulære risikofaktorer, mindre røyking og redusert eksponering for miljøgifter, men her trengs det mer forskning.
Den positive trenden er imidlertid ingen garanti for fremtiden. Nyere data tyder på at Flynn-effekten har snudd i Norge (Bratsberg & Rogeberg, 2018), altså at unge i dag skårer lavere på kognitive tester enn for 20–30 år siden. I tillegg peker internasjonale undersøkelser som PISA 2022 og studier av læringstap etter COVID-19-pandemien mot betydelige reduksjoner i faglige prestasjoner og kognitiv utvikling hos barn og unge (Jakubowski et al., 2023; OECD, 2023). Dersom lavere skoleprestasjoner og kognitive skårer i oppveksten også vil slå ut på kognitiv funksjon i aldring, kan dagens nedgang i demensrisiko snu (Figur 4). Fremtidige generasjoner kan da møte en dobbel ulempe: lavere utgangsnivå kombinert med mer ustabil kognitiv funksjon i aldring.
Figur 4
Sammenheng mellom kognitive trender og fremtidig demensrisiko
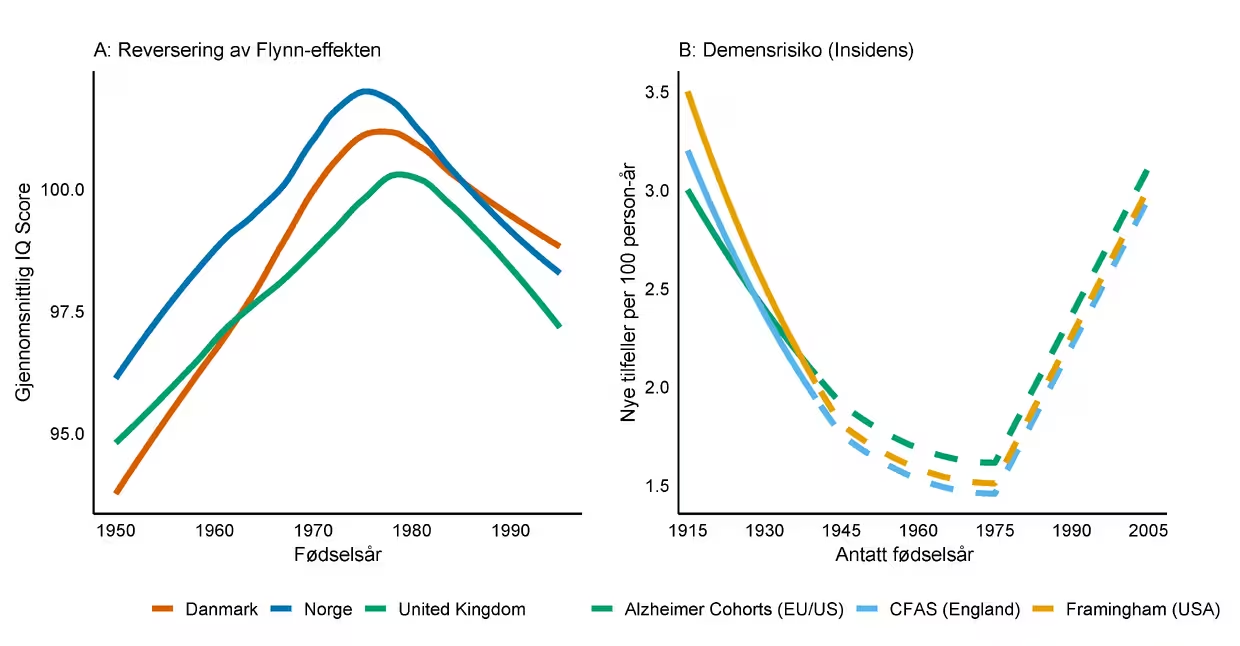
Merknad. Figuren illustrerer hvordan historiske endringer i befolkningens kognitive testresultater (Flynn-effekten) kan henge sammen med trender i forekomst av demens. Panel A viser den betydelige økningen i gjennomsnittlig IQ-skår gjennom store deler av 1900-tallet, etterfulgt av en nedgang i senere kohorter. Dette er en skjematisk fremstilling av kohortrelaterte kognitive trender, basert på rapporterte reverseringer av Flynn-effekten i Norge, Danmark og Storbritannia. Kurvene er forenklede, delvis ekstrapolerte, og ikke direkte gjengivelser av de opprinnelige dataene. Panel B illustrerer den observerte nedgangen i risikoen for å utvikle demens (insidens) for personer født tidlig på 1900-tallet. Historiske data (heltrukken linje) er basert på store internasjonale studier. De stiplede linjene representerer en spekulativ projisering av fremtidig demensrisiko gitt en hypotese om at deler av endringene vi ser, er assosiert med Flynn-effekten.
Dette er foreløpig spekulasjoner – vi vet ikke om Flynn-effekten faktisk er relatert til redusert demensforekomst, og om en reversering vil ha tilsvarende negative konsekvenser – men vi mener et slikt scenario er sannsynlig nok til at vi må ta det alvorlig. Den observerte nedgangen i demensrisiko er med andre ord ikke en naturkraft, men et resultat av gunstige samfunnsforhold som må vedlikeholdes og videreutvikles for barn og unge i dag og i fremtiden. Hvis vi ønsker å bevare, og helst forsterke, den positive trenden, må vi fortsette å investere i faktorene som bygger kognitiv kapasitet og motstandskraft gjennom hele livsløpet, og spesielt i de tidlige leveårene.
Referanser
Batsberg, B., Fjell, A. M., Rogeberg, O. J., Skirbekk, V. F. & Walhovd, K. B. (2024). Differences in cognitive function at 18 y of age explain the association between low education and early dementia risk. Proceedings of the National Academy of Sciences of the U S A, 121(41), e2 412 017 121. https://doi.org/10.1073/pnas.2412017121
Bratsberg, B. & Rogeberg, O. (2018). Flynn effect and its reversal are both environmentally caused. Proceedings of the National Academy of Sciences of the U S A, 115(26), 6674–6678. https://doi.org/10.1073/pnas.1718793115
Deary, I. J., Pattie, A. & Starr, J. M. (2013). The stability of intelligence from age 11 to age 90 years: the Lothian birth cohort of 1921. Psychological Science, 24(12), 2361–2368. https://doi.org/10.1177/0956797613486487
Engvig, A., Fjell, A. M., Westlye, L. T., Moberget, T., Sundseth, O., Larsen, V. A. & Walhovd, K. B. (2010). Effects of memory training on cortical thickness in the elderly. Neuroimage, 52(4), 1667–1676. https://doi.org/10.1016/j.neuroimage.2010.05.041
Fjell, A. M., Grydeland, H., Krogsrud, S. K., Amlien, I., Rohani, D. A., Ferschmann, L., Storsve, A. B., Tamnes, C. K., Sala-Llonch, R., Due-Tonnessen, P., Bjornerud, A., Solsnes, A. E., Haberg, A. K., Skranes, J., Bartsch, H., Chen, C. H., Thompson, W. K., Panizzon, M. S., Kremen, W. S.,…Walhovd, K. B. (2015). Development and aging of cortical thickness correspond to genetic organization patterns. Proceedings of the National Academy of Sciences of the U S A, 112(50), 15 462–15 467. https://doi.org/10.1073/pnas.1508831112
Fjell, A. M., Rogeberg, O., Sorensen, O., Amlien, I. K., Bartres-Faz, D., Brandmaier, A. M., Cattaneo, G., Duzel, S., Grydeland, H., Henson, R. N., Kuhn, S., Lindenberger, U., Lyngstad, T. H., Mowinckel, A. M., Nyberg, L., Pascual-Leone, A., Sole-Padulles, C., Sneve, M. H., Solana, J.,…Vidal-Pineiro, D. (2025). Reevaluating the role of education on cognitive decline and brain aging in longitudinal cohorts across 33 Western countries. Nature Medicine, 31(9), 2967–2976. https://doi.org/10.1038/s41591-025-03828-y
Fjell, A. M., Sorensen, O., Wang, Y., Amlien, I. K., Baare, W. F. C., Bartres-Faz, D., Bertram, L., Boraxbekk, C. J., Brandmaier, A. M., Demuth, I., Drevon, C. A., Ebmeier, K. P., Ghisletta, P., Kievit, R., Kuhn, S., Madsen, K. S., Mowinckel, A. M., Nyberg, L., Sexton, C. E.,…Walhovd, K. B. (2023). No phenotypic or genotypic evidence for a link between sleep duration and brain atrophy. Nature Humane Behaviour, 7(11), 2008–2022. https://doi.org/10.1038/s41562-023-01707-5
Fjell, A. M. & Walhovd, K. B. (2024). Individual sleep need is flexible and dynamically related to cognitive function. Nature Human Behaviour, 8(3), 422–430. https://doi.org/10.1038/s41562-024-01827-6
Grodem, E. O. S., Vidal-Pineiro, D., Sorensen, O., Bartres-Faz, D., Brandmaier, A. M., Cattaneo, G., Garrido, P. F., Henson, R. N., Kuhn, S., Lindenberger, U., MacIntosh, B. J., Nyberg, L., Pascual-Leone, A., Smith, S. M., Sole-Padulles, C., Solana-Sanchez, J., Watne, L. O., Walhovd, K. B., Bjornerud, A. & Fjell, A. M. (2025). Distinguishing Lifelong Individual Differences from Divergent Aging Trajectories of Adult Brain Volumes. bioRxiv. https://doi.org/10.1101/2025.05.26.655710
Jakubowski, M., Gajderowitcz, T. & Patrinos, H. A. (2023). Global learning loss in student achievement: First estimates using comparable reading scores. Economic Letters, 232, 111 313.
Lin, F. R., Pike, J. R., Albert, M. S., Arnold, M., Burgard, S., Chisolm, T., Couper, D., Deal, J. A., Goman, A. M., Glynn, N. W., Gmelin, T., Gravens-Mueller, L., Hayden, K. M., Huang, A. R., Knopman, D., Mitchell, C. M., Mosley, T., Pankow, J. S., Reed, N. S., …Group, A. C. R. (2023). Hearing intervention versus health education control to reduce cognitive decline in older adults with hearing loss in the USA (ACHIEVE): a multicentre, randomised controlled trial. Lancet, 402(10 404), 786–797. https://doi.org/10.1016/S0140-6736(23)01406-X
Livingston, G., Huntley, J., Liu, K. Y., Costafreda, S. G., Selbaek, G., Alladi, S., Ames, D., Banerjee, S., Burns, A., Brayne, C., Fox, N. C., Ferri, C. P., Gitlin, L. N., Howard, R., Kales, H. C., Kivimaki, M., Larson, E. B., Nakasujja, N., Rockwood, K., …Mukadam, N. (2024). Dementia prevention, intervention, and care: 2024 report of the Lancet standing Commission. Lancet, 404(10 452), 572–628. https://doi.org/10.1016/S0140-6736(24)01296-0
Ngandu, T., Lehtisalo, J., Solomon, A., Levalahti, E., Ahtiluoto, S., Antikainen, R., Backman, L., Hanninen, T., Jula, A., Laatikainen, T., Lindstrom, J., Mangialasche, F., Paajanen, T., Pajala, S., Peltonen, M., Rauramaa, R., Stigsdotter-Neely, A., Strandberg, T., Tuomilehto, J.,…Kivipelto, M. (2015). A 2 year multidomain intervention of diet, exercise, cognitive training, and vascular risk monitoring versus control to prevent cognitive decline in at-risk elderly people (FINGER): a randomised controlled trial. Lancet, 385(9984), 2255–2263. https://doi.org/10.1016/S0140-6736(15)60461-5
Nyberg, L., Lövdén, M., Riklund, K., Lindenberger, U. & Bäckman, L. (2012). Memory aging and brain maintenance. Trends in Cognitive Sciences, 16(5), 292–305. https://doi.org/10.1016/j.tics.2012.04.005
OECD. (2023). PISA 2022 Results (Volume I): The State of Learning and Equity in Education.
Pietschnig, J. & Voracek, M. (2015). One Century of Global IQ Gains: A Formal Meta-Analysis of the Flynn Effect (1909–2013). Perspective of Psychological Science, 10(3), 282–306. https://doi.org/10.1177/1745691615577701
Ritchie, S. J. & Tucker-Drob, E. M. (2018). How Much Does Education Improve Intelligence? A Meta-Analysis. Psychological Science, 29(8), 1358–1369. https://doi.org/10.1177/0956797618774253
Roe, J. M., Vidal-Pineiro, D., Sorensen, O., Grydeland, H., Leonardsen, E. H., Iakunchykova, O., Pan, M., Mowinckel, A., Stromstad, M., Nawijn, L., Milaneschi, Y., Andersson, M., Pudas, S., Brathen, A. C. S., Kransberg, J., Falch, E. S., Overbye, K., Kievit, R. A., Ebmeier, K. P., …Wang, Y. (2024). Brain change trajectories in healthy adults correlate with Alzheimer’s related genetic variation and memory decline across life. Nature Communications, 15(1), 10 651. https://doi.org/10.1038/s41467-024-53548-z
Vidal-Pineiro, D., Sorensen, O., Stromstad, M., Amlien, I. K., Baare, W., Bartres-Faz, D., Brandmaier, A. M., Cattaneo, G., Duzel, S., Ghisletta, P., Henson, R. N., Kuhn, S., Lindenberger, U., Mowinckel, A. M., Nyberg, L., Pascual-Leone, A., Roe, J. M., Solana-Sanchez, J., Sole-Padulles, C.,…Fjell, A. M. (2025). Vulnerability to memory decline in aging revealed by a mega-analysis of structural brain change. Nature Communications, 16(1), 11 488. https://doi.org/10.1038/s41467-025-66354-y
Walhovd, K. B., Krogsrud, S. K., Amlien, I. K., Bartsch, H., Bjornerud, A., Due-Tonnessen, P., Grydeland, H., Hagler, D. J., Jr., Haberg, A. K., Kremen, W. S., Ferschmann, L., Nyberg, L., Panizzon, M. S., Rohani, D. A., Skranes, J., Storsve, A. B., Solsnes, A. E., Tamnes, C. K., Thompson, W. K., …Fjell, A. M. (2016). Neurodevelopmental origins of lifespan changes in brain and cognition. Proceedings of the National Academy of Sciences of the U S A, 113(33), 9357–9362. https://doi.org/10.1073/pnas.1524259113
Walhovd, K. B., Lovden, M. & Fjell, A. M. (2023). Timing of lifespan influences on brain and cognition. Trends in Cognitive Sciences, 27(10), 901–915. https://doi.org/10.1016/j.tics.2023.07.001
Weintraub, S., Gefen, T., Geula, C. & Mesulam, M. M. (2025). The first 25 years of the Northwestern University SuperAging Program. Alzheimers Dement, 21(8), e70 312. https://doi.org/10.1002/alz.70312
Wolters, F. J., Chibnik, L. B., Waziry, R., Anderson, R., Berr, C., Beiser, A., Bis, J. C., Blacker, D., Bos, D., Brayne, C., Dartigues, J. F., Darweesh, S. K. L., Davis-Plourde, K. L., de Wolf, F., Debette, S., Dufouil, C., Fornage, M., Goudsmit, J., Grasset, L., …Hofman, A. (2020). Twenty-seven-year time trends in dementia incidence in Europe and the United States: The Alzheimer Cohorts Consortium. Neurology, 95(5), e519–e531. https://doi.org/10.1212/WNL.0000000000010022